| |
Irgendwann in einem begrenzten Zeitraum der
Embryonalentwicklung wird nach einer Mitose das eine der
beiden X-Chromosomen nicht mehr entspiralisiert. Es
verbleibt in der Transportform und ist damit inaktiviert.
Die genetische Information kann nun nicht mehr abgelesen
werden und für die Tochterzellen bleiben nur noch die
Informationen des anderen X-Chromosoms übrig. Ist bei einer
sich entwickelnden Schildpatt-Katze das X-Chromosom von der
Mutter inaktiviert, verfügen diese Zelle und alle von ihr
durch Mitose abstammenden Zellen nur noch über das Ox-Allel
auf dem X-Chromosom vom Vater. Ist dagegen das vom Vater
stammende inaktiviert, verbleibt als Informationsquelle nur
das von der Mutter mit dem ox-Allel.
Erstere sind in der Zeichnung mit O gekennzeichnet und
werden später einmal nach vielen weiteren Mitosen die
orangefarbenen Flecken mit dem weiter oben schon
beschriebenen Genotyp abgeben. Jetzt können Sie sich auch
das Fragezeichen erklären, dahinter verbirgt sich das Allel
des inaktivierten X-Chromosoms.

In der letzten Zeichnung sehen Sie, wie die Farbflächen
wachsen. Sie können leicht ableiten, daß die geschlossenen
Flächen einer Farbe größer werden, wenn die Inaktivierung
früh in der Entwicklung stattfindet und klein bleiben, wenn
die Inaktivierung spät erfolgt. Liegt der
Inaktivierungsvorgang jedoch zu früh, noch vor der
Differenzierung in die verschiedenen Organanlagen, dann ist
auch eine genetische Schildpatt-Katze einfarbig, weil ja
dann alle Hautzellen den gleichen Inaktivierungszustand
vererbt bekommen.
Vom züchterischen Standpunkt aus liegt genau da der Hase im
Pfeffer! Der Zeitpunkt der Inaktivierung des zweiten
X-Chromosoms ist weder vorhersehbar noch beeinflußbar. Und
welches der beiden X-Chromosomen inaktiviert wird, das
bleibt für jede Ausgangszelle unabhängig von den
Nachbarzellen dem ZUFALL überlassen. Weder Reinzucht noch
Linienzucht führen zum Erfolg. Trotzdem sind auf
Ausstellungen immer wieder sehr harmonisch gefärbte
Schildpatt-Katzen zu bewundern. Hut ab vor der Geduld der
Züchter!
Nachdem wir
jetzt wissen wie die Schildpatt-Zeichnung zustande kommt,
sind wir mal wieder reif, nicht für die Insel, sondern für
eine neue Tabelle. Allerdings ist es jetzt schon etwas
kompliziert, alle Möglichkeiten in einer noch einigermaßen
verständlichen Form zusammenzufassen. Die folgende Tabelle
schließt alle Farbkompositionen ein, an denen das Orange-Gen
beteiligt ist. Vorab noch ein paar Erläuterungen und
Wiederholungen zum besseren Verständnis.
Das Orange-Gen gehört zu einem System von Polygenen, das
auch als Rufus-Gruppe bezeichnet wird. Das Orange-Gen ist
das Hauptgen (engl.: major gene), das exakt den Mendel'schen
Regeln für die geschlechtsgebundene Vererbung folgt. Die
übrigen Gene dieses System verbergen sich als Untergene
(engl.: minor genes) in dem gesamten genotypischen Milieu
und beeinflussen die Ausprägung des betreffenden Merkmals
mehr oder weniger stark und fast immer in kaum berechenbarer
Weise. Die minor genes sind verantwortlich für die für
manche Merkmale charakteristische Modifikationsbreite. Rot
variiert von hellgelb bis tieforange durch diese
Modifikatoren oder minor genes. Dann dürfen wir natürlich
auch bei Orange die Modifikatoren der Dilution-Gruppe nicht
vergessen, die mit dem Verdünnungs-Allel (d) des Hauptgens
alle Schattierungen von Creme ermöglichen. Analysiert man
die Verteilung der Farbpartikel bei den verdünnten Farben
mikroskopisch, findet man deutlich unterscheidbare
Zahlenverhältnisse. Sie folgen den Mendel'schen Regeln eines
dominanten epistatischen Erbgangs gegenüber d und eines
hypostatischen Erbgangs gegenüber D. Das Gen wurde von
Patricia Turner dilution modifier (Dm) genannt und die
Ergebnisse 1980 veröffentlicht. Die mit dem unbewaffneten
Auge sichtbaren Auswirkungen mögen gegenüber der
Variationsbreite der übrigen Modifikatoren der
Dilution-Gruppe und auch der Rufus-Gruppe minimal und von
züchterischen Standpunkt irrelevant sein, für den Genetiker
ist es allemal interessant. Es kann auch sein, daß die Farbe
apricot für die modifizierte Verdünnung von orange, caramel
für die von schwarz und taupe für die von chocolate so in
den Rassestandards nicht vorkommen. Ich versuche nur, die
Farben und deren Variationen aus der Sicht des Genetikers
möglichst anschaulich zu beschreiben, die Einordnung in
Standard-Kategorien muß ich Richtern und Züchtern
überlassen.
Zurück zu unserem Thema. Nicht die Rufus-Gruppe allein ist
insbesondere für die Ausprägung der roten Farbe
ausschlaggebend, auch die Polygene der Ticking-Gruppe
spielen eine große Rolle. Diese Polygene kontrollieren die
Bänderung der Haare und nehmen damit Einfluß auf die
Intensität und Flächigkeit der Tabby-Zeichnung. Die
Modifikatoren haben auf das Classic-Allel (tb)
des Tabby-Hauptgens eine besonders ausgeprägte Wirkung. Die
dunkel-orange Zeichnung ist besonders großflächig und
überdeckt im Idealfall nahezu die gesamte hellere
Agouti-Grundfarbe. Die Orange-Flecken erscheinen dann
einfarbig. Deshalb steht in der letzten Spalte (tb/tb),
auch wenn es sich um einen Non-Agouti-Genotyp handelt, denn
die übrigen Farbflächen sollen ja einfarbig sein bei der
orginären Schildpatt- oder Tortie-Katze. Anders ist das
natürlich bei der Tortie-Tabby oder Torbie-Katze, bei der
sowohl im Orange als auch in den anderen Farbflächen eine
deutliches Tabbymuster gewünscht wird.
Ich möchte, bevor Sie die folgende Tabelle genau unter die
Lupe nehmen, noch mal darauf hinweisen, daß wir es hier
genetisch mit ein- oder zweifarbigen Katzen zu tun haben.
Orange in allen unverdünnten oder verdünnten Schattierungen
mit mehr oder weniger deutlicher Tabby-Zeichnung und
Schildpatt. Letztere sind zweifarbig. Die orangenen Bezirke
entsprechen der Beschreibung bei den Einfarbigen. Die
andersfarbigen Bezirke können einfarbig schwarz oder
einfarbig chocolate sein, wobei jeweils alle Verdünnungen
möglich sind. Schildpatt-Katzen, bei denen sowohl die
Orange-Bezirke als auch die Nichtorange-Bezirke
Tabby-Zeichnung tragen, sind Agoutis (A/-). Solche, bei
denen höchstens die Orange-Bezirke gemustert sind, sind
Non-Agoutis (a/a).
Bei den in Klammern gesetzten Allelen merken Sie sich
folgende Lesart: O/O, O/o sind verkürzte Schreibweisen von Ox/Ox
und Ox/ox.
Bei Katern ist das zweite X-Chromosom durch das y-Chromosom
zu ersetzen. Das geht nur bei den in Klammern gesetzten
Allelen, ohne daß sich am Phänotyp etwas ändert. Sie sehen
also, Kater sind immer einfarbig, rot oder red-tabby! Oder
vielleicht doch nicht? Wir werden sehen.
| |
Mögliche
Genotypen Orange |
| |
|
| |
red: |
-- |
-- |
C- |
D- |
-- |
ii |
O(O) |
tbtb |
| |
red-...-tabby: |
(A-) |
-- |
C- |
D- |
-- |
ii |
O(O) |
** |
| |
schildpatt: |
aa |
B- |
C- |
D- |
-- |
ii |
Oo |
tbtb |
| |
(black)-...-torbie: |
A- |
B- |
C- |
D- |
-- |
ii |
Oo |
** |
| |
(echter) roter Abessinier: |
-- |
-- |
C- |
D- |
-- |
ii |
O(O) |
Ta- |
| |
red-point: |
aa |
-- |
cscs |
D- |
-- |
ii |
O(O) |
tbtb |
| |
red-tabby-point: |
(A-) |
-- |
cscs |
D- |
-- |
ii |
O(O) |
** |
| |
seal-tortie-point: |
aa |
B- |
cscs |
D- |
-- |
ii |
Oo |
tbtb |
| |
seal-torbie-point: |
A- |
B- |
cscs |
D- |
-- |
ii |
Oo |
** |
| |
cameo-...-tabby |
(A-) |
-- |
C- |
D- |
-- |
I- |
O(O) |
** |
| |
red-smoke, -shaded, -shell: |
-- |
-- |
C- |
D- |
-- |
I- |
O(O) |
tbtb |
| |
cameo-...-torbie: |
A- |
B- |
C- |
D- |
-- |
I- |
Oo |
** |
| |
schildpatt-smoke, -shaded, -shell: |
aa |
B- |
C- |
D- |
-- |
I- |
Oo |
tbtb |
| |
|
|
|
|
|
|
|
|
|
| |
creme: |
-- |
-- |
C- |
dd |
dmdm |
ii |
O(O) |
tbtb |
| |
apricot: |
-- |
-- |
C- |
dd |
Dm- |
ii |
O(O) |
tbtb |
| |
creme-...-tabby |
(A-) |
-- |
C- |
dd |
dmdm |
ii |
O(O) |
** |
| |
blau-creme (-schildpatt): |
aa |
B- |
C- |
dd |
dmdm |
ii |
Oo |
tbtb |
| |
(blau-)creme-...-torbie: |
A- |
B- |
C- |
dd |
dmdm |
ii |
Oo |
** |
| |
(echter) creme Abessinier: |
-- |
-- |
C- |
dd |
dmdm |
ii |
O(O) |
Ta- |
| |
creme-point: |
aa |
-- |
cscs |
dd |
dmdm |
ii |
O(O) |
tbtb |
| |
creme-tabby-point: |
(A-) |
-- |
cscs |
dd |
dmdm |
ii |
O(O) |
** |
| |
blue-tortie-point: |
aa |
B- |
cscs |
dd |
dmdm |
ii |
Oo |
tbtb |
| |
blue-torbie-point: |
A- |
B- |
cscs |
dd |
dmdm |
ii |
Oo |
** |
| |
cameo-creme-...-tabby: |
(A-) |
-- |
C- |
dd |
dmdm |
I- |
O(O) |
** |
| |
creme-smoke, -shaded, -shell: |
-- |
-- |
C- |
dd |
dmdm |
I- |
O(O) |
tbtb |
| |
blaucreme-smoke, -shaded, -shell: |
aa |
B- |
C- |
dd |
dmdm |
I- |
Oo |
tbtb |
| |
|
|
|
|
|
|
|
|
|
| |
chocolate-schildpatt: |
aa |
b* |
C- |
D- |
-- |
ii |
Oo |
tbtb |
| |
chocolate-...-torbie: |
A- |
b* |
C- |
D- |
-- |
ii |
Oo |
** |
| |
chocolate-tortie-point: |
aa |
bb |
cscs |
D- |
-- |
ii |
Oo |
tbtb |
| |
chocolate-torbie-point: |
A- |
bb |
cscs |
D- |
-- |
ii |
Oo |
** |
| |
lilac-schildpatt: |
aa |
b* |
C- |
dd |
dmdm |
ii |
Oo |
tbtb |
| |
lilac-...-torbie: |
A- |
b* |
C- |
dd |
dmdm |
ii |
Oo |
** |
| |
lilac-tortie-point: |
aa |
bb |
cscs |
dd |
dmdm |
ii |
Oo |
tbtb |
| |
lilac-torbie-point: |
A- |
bb |
cscs |
dd |
dmdm |
ii |
Oo |
** |
| |
cinnamon-schildpatt: |
aa |
bl/bl |
C- |
D- |
-- |
ii |
Oo |
tbtb |
| |
fawn-schildpatt: |
aa |
bl/bl |
C- |
dd |
dmdm |
ii |
Oo |
tbtb |
| |
burma-rot: |
aa |
-- |
cbcb |
D- |
-- |
ii |
O(O) |
-- |
| |
burma-creme: |
aa |
-- |
cbcb |
dd |
dmdm |
ii |
O(O) |
-- |
| |
|
|
|
|
|
|
|
|
|
| |
Anmerkungen: |
|
|
|
|
|
|
|
|
| |
* = bei "mackerel"
(TT) oder (Ttb)
, bei
"classic" (tbtb)
|
|
| |
... = je
nach Genotyp durch "mackerel" oder "classic" zu
ersetzen, wie bei black-...- tabby gezeigt. |
|
| |
b* =
bb oder bbl |
|
| |
tortie
= schildpatt |
|
| |
torbie
= tortie-tabby |
|
| |
cameo
= red-silver |
|
| |
shell
= red-chinchilla |
|
| |
(A-) =
auch Non-Agoutis sind gezeichnet, aber bei Agoutis
ist die Tabby-Zeichnung noch deutlicher |
Schildpatt-Kater
Ich kann es nicht länger ignorieren, es gibt sie doch.
Schildpatt-Kater sind allerdings sehr selten, sel-ten
zeugungsfähig und haben häufig eine kürzere Lebenserwartung.
Wenn sie denn doch einen Wurf zustande bringen, ist das
Ergebnis fast immer frustrierend. Auf jeden Fall ist ein
Verpaarung einer Schilpatt-Katze mit einem der seltenen
Schilpatt-Kater keine Grundlage zu einer
Schildpatt-Reinzucht. Um das zu verstehen, müssen wir ganz
weit zurückgehen, sozusagen an den Anfang, nicht der Welt,
sondern dieses Artikels. Meiose heißt das Thema.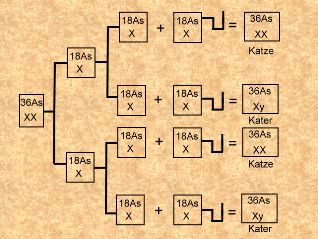
Normale Meiose und Befruchtung
mit normalen Spermien (Bild rechts):
Zur Wiederholung: In den
Ovarien sind die Vorläufer der Eizellen oder Oocyten, die
Oogonien. In der ersten meiotischen Teilung wird der
diploide oder doppelte auf den haploiden oder einfachen
Chromosomensatz reduziert, weshalb die ganze Meiose auch
Reduktionsteilung heißt. Aber die Reduktion findet
tatsächlich nur im ersten Teil der Meiose statt. Der
diploide Satz hat 38 Chromosomen, 36 Autosomen (As) und die
2 Geschlechtschromosomen (XX), jedes Chromosom besteht aus
zwei Chromatiden. Die beiden Tochterzellen aus der ersten
meiotischen Teilung haben jeweils einen vollständigen
haploiden Chromosomensatz (18As + X), aber jedes Chromosom
besteht nach wie vor aus zwei Chromatiden. Erst in der
zweiten meiotischen Teilung werden die Chromatiden verteilt,
ähnlich wie bei der Mitose. Das Ergebnis sind vier
Teilungsprodukte mit gleicher Gen- aber eventuell
unterschiedlicher Allelenausstattung. Jede der vier Zellen
hat die gleiche Chance, sich zur Eizelle zu entwickeln. Der
Zufall allein entscheidet, welche tatsächlich zur Eizelle
wird. Die restlichen drei Zellen werden zu Polkörpern und
haben nicht unwichtige Aufgaben bei der Bildung des
Follikels, in dem die Eizelle heranreift und bis zum
Eisprung verbleibt. Und weil jedes der vier Teilungsprodukte
die gleiche Chance zur Eizelle hat, müssen wir bei einer
Analyse auch alle vier Möglichkeiten in Betracht ziehen.
Deshalb sind in der Schemazeichnung auch alle vier möglichen
Eizellen aufgeführt, tatsächlich überlebt aus einer
einzelnen Meiose natürlich nur eine als Eizelle. Bitte das
nicht vergessen und nicht verwechseln.
Beim Kater verläuft die Meiose in Prinzip gleich, nur daß
sich alle vier Teilungsprodukte zu Spermien entwickeln.
Außerdem entstehen durch die Reduktion in der ersten
meiotischen Teilung zwei unterschiedliche Typen von
Spermien. Der Kater verfügt ja über zwei verschiedene
Geschlechtschromosomen, nämlich ein X-Chromosom und ein
y-Chromosom. Dementsprechend bekommt nach der ersten Teilung
die eine Zelle das X-Chromosom und die andere das
y-Chromosom, jeweils natürlich zusätzlich zum haploiden
Autosomensatz. Nach der zweiten Teilung haben wir dann vier
Teilungsprodukte und alle vier entwickeln sich zu Spermien.
Dabei sind jeweils zwei Spermien identisch und werden
weiblich bestimmend genannt, wenn sie das X-Chromosom haben
und männlich bestimmend, wenn sie das y-Chromosom abbekommen
haben.
Der entscheidende Unterschied zwischen der Meiose beim Kater
und der bei der Katze liegt in den zeitlichen Abläufen. Beim
Kater finden in den Hodengeweben dauernd Meiosen statt. Sie
dauern nur wenige Stunden und die Entwicklung zu fertigen
Spermien geht rasch voran. Sie werden eine gewisse Zeit
gespeichert und wenn es nicht zum Deckakt kommt, dann werden
sie wieder abgebaut und vom Körper aufgenommen, denn sie
enthalten ja eine Menge wertvoller Substanzen, die der
Körper wieder verwerten kann. Die Spermien sind bei der
Befruchtung der aktive Partner. Sie müssen nach der
Ejakulation durch den Uterus zum Eileiter schwimmen und
treffen erst dort auf die befruchtungsfähige Eizelle - ein
gewaltiger Weg für so ein kleines Gebilde. Trägt ein
Spermium irgendeine Störung, sei es ein Chromosom zuviel
oder zuwenig, sei es eine ungünstige Allelenkombination,
dann wird es sich erst gar nicht entwickeln oder es ist
bewegungsunfähig oder sehr viel langsamer als die anderen
Spermien. Daher liegt es fast nie am Kater, wenn es in der
befruchteten Eizelle zu unkonventionellen
Chromosomenkonstellationen kommt, weil in der Regel nur
"normale" Spermien bis zur Eizelle vordringen.
Bei der Katze sieht die ganze Sache erheblich anders aus.
Wir müssen drei verschiedene Zyklen unterscheiden:
1) der anovulatorische Zyklus dauert 14-28 Tage
2) der ovulatorische Zyklus dauert 14-16 Wochen, nämlich ca.
9 Wochen für die Trächtigkeit und ca. 6 Wochen für die
Säugeperiode
3) der pseudogravide Zyklus mit 40-50 Tagen
Alle drei Zyklen beginnen gleich. Einige oder einige zehn
Oogonien beginnen mit der Meiose und der Follikelbildung.
Nachdem sich die homologen Chromosomen in der Prophase-I
vollständig gepaart haben, also Bivalente gebildet haben,
ist erst einmal eine Pause, die Katze ist jetzt rollig. Wird
sie nicht gedeckt (anovulatorischer Zyklus) werden innerhalb
von 14-28 Tagen die in der Metaphase-I steckengebliebenen
Oocyten abgebaut und resorbiert und das ganze Spiel beginnt
von vorn. Bleiben die Follikel erhalten, ist die Katze
dauerrollig. Platzen die Follikel ohne daß eine Deckung und
nachfolgende Befruchtung stattgefunden hat, kommt es zur
Scheinschwangerschaft (Pseudogravidität). Die Oocyten werden
zwar resorbiert, die Hormone aus den geplatzten Follikeln
täuschen jedoch eine Schwangerschaft vor. Wird die Katze
jedoch am 2.-5. Tag der Rolligkeit gedeckt, werden die
Oocyten durch den Deckakt angeregt, mit Meiose
weiterzumachen und sie innerhalb von 24 Stunden
abzuschließen. Inzwischen hat sich auch der Follikel voll
entwickelt, nicht zuletzt mit Hilfe der Polzellen. Jetzt und
nach jedem weiteren Deckakt platzt ein Follikel und die
Eizellen wandern zum Eileiter und werden dort von den
Spermien befruchtet. Inzwischen haben die Follikelhormone
den Körper auf die Schwangerschaft vorbereitet und dafür
gesorgt, daß sich die befruchteten Eizellen in der
Gebärmutter einnisten können (ovulatorischer Zyklus).
Zeit spielt hier die entscheidende Rolle. Die Oocyten
verharren ein bis mehrere Tage in der Prophase-I der Meiose,
die Homologen sind vollständig zu Bivalenten gepaart. Es
kann zu mehrfachen Überkreuzungen der vier beteiligten
Chromatiden und zu dauerhaften Verklebungen kommen. In der
folgen Anaphase-I können sich dann die homologen Chromosomen
nicht oder nicht mehr sauber trennen (Nondisjunction) und es
kommt zu Fehlverteilungen in den beiden entstehenden
Tochterzellen. Die zweite meiotische Teilung verläuft in den
meisten Fällen normal, hier werden ja Chromatiden
voneinander getrennt. Aber das nützt dann auch nichts mehr,
die in der Anaphase-I begonnene Fehlverteilung bleibt auch
den Teilungsprodukten der Anaphase-II erhalten. Ist ein
Autosomenbivalent von einem Nondisjunction betroffen, geht
entweder schon die Eizelle selbst oder dann die befruchtete
Eizelle (Zygote) wegen der extrem unausgewogenen
Gendosiswirkung zugrunde. Gerät die Genbalance nicht so
stark aus dem Gleichgewicht, weil eines der kleineren
Autosomenbivalente beteiligt ist, entwickelt sich oft noch
ein Embryo, der dann aber in einem sehr frühen Stadium
abgestoßen und resorbiert wird. Wir brauchen uns also mit
derartigen Chromosomenstörungen oder -aberrationen nicht
weiter aufzuhalten.
Ganz anders verhält es sich, wenn das X-Chromosomenbivalent
ein Nondisjunction durchmacht. Hier kann die gestörte
Gendosiswirkung durch Inaktivierung entsprechend der
Lyon-Hypothese kompensiert werden. Dabei kann auch mehr als
ein X-Chromosom inaktiviert werden und der Embryo entwickelt
sich dann nahezu normal. Selbst Jungtiere erscheinen normal,
erst später zeigen sich Störungen, weil die Inaktivierung
der X-Chromosomen eben doch nicht ganz vollständig ist und
weil ja das Erbmaterial trotzdem vorhanden ist und die
Genbalance stört. Aber schauen wir uns erst einmal in Ruhe
das nachfolgende Schema mit einem Nondisjunction der
X-Chromosomen an und vergleichen es mit dem ersten Schema
einer normalen Meiose.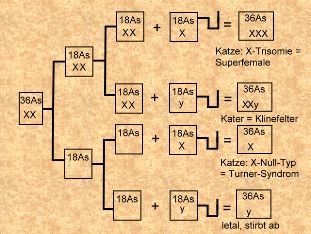 Nondisjunktion in der Meiose I und Befruchtung mit normalen
Spermien (Bild links).
Nondisjunktion in der Meiose I und Befruchtung mit normalen
Spermien (Bild links).
Wie sie sehen, kommt es zu
einer ganzen Reihe von ungewöhnlichen Konstellationen. Man
kann den Typ der Aberration übrigens sehr gut mit einer
einfachen Untersuchung von Körper- oder Blutzellen
nachweisen. Wir haben früher schon festgestellt, daß die
Chromosomen im Interphasekern, also dem Zustand, der
zwischen zwei Teilungen liegt und die eigentliche
Arbeitsform darstellt, nicht sichtbar sind, weil sie maximal
entspiralisiert sind. Aber gerade im Interphasekern heben
sich die Inaktivierten, also nicht entspiralisierten
X-Chromosomen als kompakte, gut anfärbbare Gebilde hervor.
Ein Forscher namens Barr hat diese Zusammenhänge 1949
erkannt und die inaktivierten X-Chromosomen werden seither
als Barr-Körperchen bezeichnet. Normale weibliche Zellen
besitzen demnach ein Barr-Körperchen, normale männliche
Zellen keines (siehe erstes Schema). Davon abweichend haben
die Katzen aus dem zweiten Schema entweder zwei oder kein
Barr-Körperchen, der Kater hat eines.
Bei allen Katzen, die sich aus Eizellen mit einem
Nondisjunction in der Meiose entwickeln, kann es sich
natürlich nicht um normale Tiere handeln. Es folgt eine
kurze Typisierung der Störungen, die sich aus diesen
ungewöhnlichen Heterosomenkombinationen ableiten lassen. Da
dieselben Probleme beim Menschen gar nicht so ungewöhnlich
sind, werden auch die Namen der Aberrationen von der
Humangenetik abgeleitet.
Die Superfemales
sind keine "Superweibchen", sondern wegen der nicht ganz
ausbalancierten Gendosiswirkung Katzen mit erheblich
gestörter Sexualentwicklung. Meistens bleibt die Rolligkeit
ganz aus und kann auch nicht medikamentös oder hormonell
hervorgerufen werden. Sollte es doch zu einer Befruchtung
kommen, sterben die Embryos sehr früh ab. Es werden nämlich
kaum normale Eizellen gebildet, weil die Homologenpaarung in
der Prophase-I der Meiose durch die X-Trisomie nahezu
unmöglich ist, was auch die Bildung der autosomalen
Bivalente ungünstig beeinflußt.
Das Turner-Syndrom
ist nach seinem Entdecker benannt und zeigt, daß die
Anwesenheit eines zweiten X-Chromosoms nicht ohne Bedeutung
ist, auch wenn es in inaktivierter Form vorliegt. Der
X-Null-Typ führt zu einem ganzen Bündel von Mißbildungen.
Die Ovarien sind unterentwickelt und in ihrer Struktur so
verändert, daß kaum Eizellen gebildet werden können. Sollte
es doch zu einer Meiose kommen, gilt auch für die
X-Monosomie, daß die Homologenpaarung erheblich gestört ist.
Auch körperlich sind solche Katzen unterentwickelt, was
bereits bei der Geburt als "Zwergenwuchs" zu erkennen ist.
Das feline Klinefelter-Syndrom
ist der Geschlechtschromosomenaberrations-Typ hinter dem
sich der Schildpatt-Kater versteckt. Einerseits sind zwei
X-Chromosomen vorhanden, die mit Hilfe des
Lyon-Inaktivierungsmechnismus Schildpatt hervorbringen.
Andererseits ist ein y-Chromosom vorhanden, das immer das
männliche Geschlecht verursacht, egal wieviel X-Chromosomen
dem entgegenstehen. Leider ist es geradezu typisch für
solche Kater, daß ihre körperlichen Entwicklung stark
verzögert abläuft und daß ihre sekundären
Geschlechtsmerkmale immer unterentwickelt bleiben.
Kleinhodigkeit ist eines der Leitmerkmale für das feline
Klinefelter-Syndrom. Wegen der gestörten Homologenpaarung
sind die Kater steril, aber immerhin noch sexuell aktiv. Nur
bringt es leider nichts.
Bis zum nächstem Mal können Sie sich ja mal überlegen, wie
denn die besprochenen Aberrationstypen hinsichtlich des
Orange-Gens aussehen können. Und vielleicht finden wir dann
auch einen Weg, wie wir doch noch zu einem fruchtbaren
Schildpatt-Kater kommen. Dabei werden wir nebenbei den Grund
dafür finden, daß eine nicht unerhebliche Zahl von
Schildpattkatzen unfruchtbar ist oder ein gestörtes
Sexualverhalten an den Tag legt.
mit freundlicher Genehmigung des Autors
Dipl. Biologe R.
Fahlisch
"Dreamhunter Cattery"
Das Copyright für den oben genannten Text, liegt sowohl beim
Autor des Textes Herrn R. Fahlisch, sowie bei dem Betreiber
dieser Seiten, Frau Ute Kunze. Eine Vervielfältigung oder
Verwendung des Textes in anderen elektronischen oder
gedruckten Publikationen ist ohne ausdrückliche Zustimmung
von Herrn Fahlisch und Frau Kunze nicht gestattet.
|
|