| |
Heute werden wir ohne besondere Einleitung die Meiose zu
Ende führen. Später fassen wir dann all das zusammen, was an
dieser besonderen Zellteilung für unser Vorhaben wichtig
ist. In der letzten Abbildung aus dem 3. Teil (Abb. 5) war
die Anaphase-I zu sehen. In der Telophase-I wandern die
beiden Chromosomengruppen zu den beiden Zellpolen und
verharren dort kurz in der Interkinese, einer speziellen
Interphase. Dabei werden die Chromosomen jedoch nicht
entspiralisiert, sondern verbleiben in der Transportform,
denn es folgt nun unmittelbar die Meiose-II. Ohne daß eine
Zellwand eingebaut oder Kernmembranen gebildet werden,
verhalten sich die beiden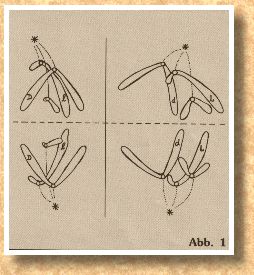 Chromosomengruppen wie zwei getrennte Zellen, die nun
synchron eine mitoseartige Teilung vollziehen. Nach einer
kurzen Prophase-II ordnen sich die beiden Chromosomengruppen
in der Metaphase-II in zwei getrennte Aquatorialplatten an.
Die beiden Teilungsebenen stehen senkrecht zu der ersten
Teilungsebene, wie wir aus Abb. l entnehmen können.
Chromosomengruppen wie zwei getrennte Zellen, die nun
synchron eine mitoseartige Teilung vollziehen. Nach einer
kurzen Prophase-II ordnen sich die beiden Chromosomengruppen
in der Metaphase-II in zwei getrennte Aquatorialplatten an.
Die beiden Teilungsebenen stehen senkrecht zu der ersten
Teilungsebene, wie wir aus Abb. l entnehmen können.
Aus den Gameten werden Spermien
Wir sehen hier die Anaphase-II, und es sieht so aus, als
ob zwei mitotische Anaphasen nebeneinander gezeichnet wären.
Die beiden identischen Chromatiden eines jeden Chromosoms
sind voneinander getrennt worden. Der kleine Unterschied
besteht darin, daß in jeder Chromosomengruppe nur noch eines
der beiden Homoigen vorhanden ist. Es geht nun ganz normal
weiter. In der Telophase-II wandern die Chromosomen
endgültig zu den vier Zellpolen und beginnen sich zu
entspiralisieren. Es werden senkrecht zueinander zwei
Zellwände eingezogen und um jede der vier Chromosomengruppen
eine Kernmembran gebildet. Das Endprodukt sind vier
voneinander unabhängige Zellen in der Interphase, die vier
Gameten, die sich in unserem Fall getrennt voneinander zu
Spermien entwickeln, denn wir haben die Meiose ja an einem
Modell-Kater betrachtet. Hätten wir unsere Katze aus dem 2.
Teil als Modell genommen, dann wäre aus einem der vier
Teilungsprodukte eine Eizelle oder Oocyte entstanden, die
übrigen drei wären zu den sog. Polkörpern verkümmert.
Welches von den vier Teilungsprodukten nun zur Eizelle wird,
bleibt allerdings dem schon so oft erwähnten ZUFALL
überlassen. Bevor wir uns aber weiter mit dem ZUFALL
befassen, wollen wir die für unsere genetischen Analysen und
Berechnungen wichtigen Schritte der Meiose noch einmal
wiederholen und zusammenfassen.
Das Genom ist die Summe aller Gene
Jeder Organismus hat einen bestimmten Bauplan, dieser
wird in Form von Genen festgehalten. Die Summe aller Gene
ist das Genom, das auf eine bestimmte Anzahl von Chromosomen
verteilt ist. Diese charakteristische Anzahl von Chromosomen
nennt man auch den einfachen oder haploiden Chromosomensatz.
Von jedem Chromosom gibt es ein Duplikat. Es ist also in
jeder Zelle ein zweiter Chromosomensatz vorhanden, der dem
ersten Satz homolog ist, lediglich die Schalterstellungen
oder Allele der entsprechenden Gene können unterschiedlich
sein. Diesen doppelten Chromosomensatz bezeichnet man als
diploiden Chromosomensatz. Dieses „doppelte System" hat sich
in der Evolution bewährt, denn bei Ausfall eines
Bauplanteils kann auf das Duplikat zurückgegriffen werden.
Damit ist ein Überleben eher gesichert. Das Vorhandensein
verschiedener Allele läßt eine größere Vielfalt zu, was
letztendlich eine Anpassung an bestimmte oder auch
veränderte Umweltbedingungen erleichtert. Daß der Mensch
diese Allelenvielfalt dazu benutzt, verschiedene Varietäten
zu züchten, ist sein Problem. Wir als Züchter müssen uns
halt notgedrungen mit der Vererbungslehre herumschlagen, um
zu einem bestimmten Ziel zu kommen. Daß uns der ZUFALL
genügend Steine auf den Weg legt, sollten wir als „Laune der
Natur" betrachten. Arrangieren müssen wir uns damit auf
jeden Fall.
Vermehrungsteilungen lassen neues Tier entstehen
In der Meiose wird der diploide Chromosomensatz auf den
haploiden reduziert, daher auch der Name Reduktionsteilung.
Dieser Schritt ist notwendig, denn durch die Verschmelzung
von zwei haploiden Gameten, der Eizelle und dem Spermium,
soll bei der Befruchtung der Grundstein für einen neuen
diploiden Organismus gelegt werden. Das
Verschmelzungsprodukt ist die befruchtete Eizelle, die
Zygote, aus der durch Vermehrungsteilungen (Mitosen) ein
neues Tier heranwächst. Hinter dem System der Meiose
verbirgt sich aber noch ein anderer interessanter Aspekt:
Durch die zufällige Anordnung des Bivalente in der
Metaphase-I werden in dem eigentlichen Reduktionsschritt,
der Anaphase-I durch die zufällige Verteilung der Homologen
auf die beiden Zellpole die Allele neu kombiniert. Die
Meiose-II ist lediglich eine mitoseartige
Vermehrungsteilung, also aus zwei mach vier, denn von vier
Gameten sind immer je zwei absolut identisch.
Kombinationsmöglichkeiten
sind einfach darzustellen
Aber schauen wir uns jetzt
die im vorigen Absatz mehrfach erwähnten ZUFÄLLE bei der
Neukombination der Allele im Verlauf der Meiose-I nochmal
genauer an. Nehmen wir unseren Modell-Kater aus dem 3. Teil
und entwickeln daraus ein einfaches Verfahren, die
Kombinationsmöglichkeiten der Allele ohne die komplizierte
Malerei von schematischen Chromosomenbildchen voraussagen zu
können. Unser Ausgangspunkt ist der in Abb. 1/3. Teil
gezeigte Chromosomensatz. Wir numerieren die
Chromosomenpaare einfach durch. Das Chromosomenpaar Nr. l
trägt das Gen für die Farbdichte mit den Allelen D/d, das
Paar Nr. 2 das Gen für die Haarlänge mit den Allelen L/l
und das Paar Nr. 3 ist das Heterosomenpaar, von dem wir
stark vereinfacht sagen können, das X-Chromosom trägt die
Schalterstellung weiblich und das Y-Chromosom die
Schalterstellung männlich. Machen wir es uns noch
einfacher und schreiben den Ge-
notyp wie folgt auf: D/d, L/l, X/Y. Diese
Schreibweise besagt, daß es sich um einen Kater (Y) handelt.
Die Fellfarbe ist unverdünnt (D), also ist das Tier bei
schwarzer Grundfarbe wirklich schwarz. Außerdem ist unser
Kater wunderschön kurzhaarig (L). Bezüglich der rezessiven
Allele d und l können wir nur spekulieren, es
sei denn, ein Eiter (Ja, Sie haben richtig gelesen! In der
genetischen Ausdrucksweise gibt es eine Einzahl von Eltern,
nämlich Elter.) unseres Katers sei blau (d/d) und
langhaarig (l/l) gewesen. Aber spekulieren wir für
unser Modell nicht, sondern nehmen den oben aufgeschriebenen
Genotyp einfach mal so hin und überprüfen später unsere
Annahme. Nehmen wir jetzt die Abb. 4/Teil 3 zu Hilfe
und spielen alle möglichen Anordnungsvarianten durch. Der
Ausgangsgenotyp unseres Modell-Katers ist immer der gleiche,
aber wenn wir die Anordnung der Bivalente in der Metaphase-I
verändern, dann ergeben sich für die Teilungsprodukte der
Anaphase-I (Abb. 5/Teil 3) eine ganze Reihe
verschiedener Allelenkombinationen.
Acht Möglichkeiten bei nur 3
Merkmalen
1. Möglichkeit:
Die Äquatorialplatte ist genau so wie aufgezeichnet. Die
beiden Spermientypen haben die Allelenkombinationen D, l, Y
und d, L, X.
2. Möglichkeit:
Drehen wir in Gedanken das l. Bivalent (l. Chromosomenpaar)
um, dann bekommen die beiden Spermientypen die Kombinationen
d, l, Y und D, L, X.
3. Möglichkeit:
Wenn wir nur das Heterosomenbivalent umdrehen und die beiden
anderen Bivalente so lassen, wie aufgezeichnet, dann führt
das zu zwei neuen Spermientypen: D, l, X und d. L, Y.
4. Möglichkeit:
Es bleibt als letzte Variante, das 2. Bivalent (2.
Chromosomenpaar) umzudrehen, die beiden anderen Bivalente
bleiben wie aufgezeichnet. Die beiden möglichen
Spermientypen sind dann D, L, Y und d, l, X.
Sortieren wir unser
Ergebnis:
D, L, X D, L, Y
D, l, X D, l, Y
d, L, X d, L, Y
d, l, X d, l, Y
Betrachten wir also für
eine Kreuzungsanalyse inclusive der Geschlechtsbestimmung
drei Merkmale, und liegen die drei Merkmale auf
verschiedenen Chromosomen, und sind alle drei Merkmale
gentypisch heterozygot, dann sind genau acht verschiedene
Typen von Spermien oder acht verschiedene
Allelenkombinationen möglich.
Drücken wir das Ganze
mathematisch aus:
Wir verfolgen die Ausprägung von n = 3 Merkmalen, die
unabhängig voneinander vererbt werden, weil die Gene auf
verschiedenen Chromosomen lokalisiert sind. Wir wissen auch,
daß von jedem Gen zwei unterschiedliche Allele vorhanden
sind, wobei wir die Geschlechtsbestimmung der Einfachheit
halber als Gen mit den Allelen weiblich (X) und männlich (Y)
betrachten. Dann können wir folgende Rechnung aufmachen:
Anzahl der
Kombinationsmöglichkeiten = 2 hoch n
In unserem speziellen Fall
sind also 2 3 = 8 Kombinationen oder Spermientypen möglich,
also genau die Anzahl, die wir auf umständliche Art weiter
oben auch schon herausbekommen haben. Übertragen wir diesen
Rechenweg auf einen richtigen Kater und berechnen nur die
Möglichkeiten, die sich aus der zufälligen Anordnung der
n = 19 Bivalente in der Metaphase-I ergeben, dann kommen
wir auf mehr als 1/2 Million Spermientypen, denn 2 hoch
19 = 524.288. Dieselbe Zahl ergibt sich natürlich auch
bei der Katze bezüglich der unterschiedlichen Eizellentypen,
denn das zweimal vorkommende X-Chromosom darf nicht
vernachlässigt werden. Es ist zwar bezüglich der
Schalterstellung „weiblich" homozygot, trägt aber noch eine
ganze Reihe anderer Gene, von denen mindestens eines
heterozygot ist, so daß die beiden X-Chromosomen mindestens
bei einem Gen unterschiedliche Merkmalsausprägungen vererben
und somit genetisch unterscheidbar sind. Aber bleiben wir
auf dem Boden der Tatsachen und verlassen das theoretische
Spiel mit den großen Zahlen. Schauen wir uns lieber eine
brauchbare Tabelle an, mit deren Hilfe wir die
Allelenkombinationen ohne die lästige Chromosomenzeichnerei
bestimmen können.
|
Allele
|
mögliche Altelenkombinationen |
|
D/d
|
D
|
d
|
|
L/l
|
L
|
1(1)
|
L
|
1(1)
|
|
X/Y
|
X
|
Y
|
X
|
Y
|
X
|
Y
|
X
|
Y
|
|
Gameten |
D
|
D
|
D
|
D
|
d
|
d
|
d
|
d
|
|
L
|
L
|
l
|
l
|
L
|
L
|
l
|
l
|
|
X.
|
Y
|
X
|
Y
|
X
|
Y
|
X
|
Y
|
In der ersten Reihe wird
das erste Allenpaar D/d untergebracht. Die zweite
Reihe enthält das zweite Paar L/l zweimal, denn jedes
Allel aus der zweiten Reihe kann ja mit jedem Allel aus der
ersten Rei-
he Kombiniert werden. In der dritten Reihe steht das dritte
Allelenpaar (hier die „Schalterstellungen"
weiblich oder männlich) X/Y vierfach, denn jedes der
beiden Allele kann ja mit den schon vorhandenen
Kombinationen aus der zweiten Reihe kombiniert werden.
Dieses System ließe sich noch beliebig fortsetzen.
Dann kommen wir aber sehr schnell wieder in den Bereich der
großen Zahlen, mit denen eine vernünftige Arbeit nicht
möglich ist. Die vierte Reihe müßte nämlich das Allenpaar
achtfach und die fünfte Reihe sechzehnfach enthalten. Wenden
wir die Tabelle auf unsere Modell-Katze vom 2. Teil an. Sie
erinnern sich sicher daran, daß es sich um eine blaue
Kurzhaar-Katze mit dem Genotyp
d/d. L/l, X/X handelt. In der ersten Reihe steht
wieder, analog zum Kater, das Allelenpaar d/d des
Verdünnungsgens.
Da aber unsere Modell-Katze für dieses Gen homozygot ist,
muß das Allel nur einmal eingetragen werden, denn das zweite
Allel ist identisch und eröffnet keine neuen
Kombinationsmöglichkeiten.
|
Allele |
mögliche Allelenkombinationen |
|
d/d
|
d
|
(d)
|
|
L/l
|
L
|
|
|
(L)
|
(l)
|
|
X/X
|
X
|
(X)
|
X
|
(X)
|
(X)
|
(X)
|
(X)
|
(X)
|
|
Gameten |
d
|
|
d
|
|
|
|
|
|
| |
L
|
|
i
|
|
|
|
|
|
| |
X
|
|
X
|
|
|
|
|
|
Ich habe nur der
Vollständigkeit halber die überflüssigen Allele in Klammern
eingetragen. In Zukunft kann man diese der Übersichtlichkeit
wegen weglassen. In der zweiten Reihe steht das Allenpaar
L/l, aber nur einmal, denn jedes der beiden Allele kann
ja nur mit dem einen d kombiniert werden. In der
dritten Reihe befindet sich schließlich das homozygote
Heterosomenpaar X/X. Auch dieses Paar muß nur einmal
auftauchen, weil die restlichen Kombinationen überflüssig
sind. Wir verfolgen auch hier drei Merkmale, aber nur n =
l Merkmale weisen zwei unterschiedliche Allele auf. Also
gibt es nur 2 hoch n = 2 hoch 1 = 2 unterschiedliche
Eizellentypen mit den Allelen d/L/X und d/l/X.
Sie sehen also, daß sowohl bei der Bildung der Spermien als
auch bei der der Eizellen schon eine ganze Menge Zufall im
Spiel ist. Aber ganz unkalkulierbar ist die Sache doch
nicht, insbesondere wenn man nur wenige Merkmale aufs Korn
nimmt. Das nächste Mal werden wir unsere beiden Modell-Tiere
verpaaren und berechnen, mit welcher Wahrscheinlichkeit wir
welches „Zucht"-Ziel erreichen können.
mit freundlicher Genehmigung des Autors
Dipl. Biologe R.
Fahlisch
"Dreamhunter Cattery"
Das Copyright für den oben genannten Text, liegt sowohl beim
Autor des Textes Herrn R. Fahlisch, sowie bei dem Betreiber
dieser Seiten, Frau Ute Kunze. Eine Vervielfältigung oder
Verwendung des Textes in anderen elektronischen oder
gedruckten Publikationen ist ohne ausdrückliche Zustimmung
von Herrn Fahlisch und Frau Kunze nicht gestattet. |
|