| |
Erinnern wir uns: Die Erbinformation ist in
Form des genetischen Codes, also in einer bestimmten
Reihenfolge von Nucleotiden, in der DNS gespeichert. Die
Information für ein bestimmtes Merkmal nennt man Gen. Ein
DNS-Molekül ist ein langes perlschnurartiges Gebilde, auf
dem viele hundert Gene hintereinander angeordnet sind. Ein
DNS-Molekül bildet zusammen mit gewissen Proteinen jeweils
ein Chromosom. Jeder höhere Organismus hat mehrere
verschiedene Chromosomen, die Summe aller darin enthaltenen
Erbinformationen nennt man Genom.
Erbinformationen werden
kopiert
Die Chromosomen befinden sich immer im
Zellkern, die Verarbeitung der Informationen und damit der
Stoffwechsel erfolgen im Zellplasma. Sowohl die Zelle nach
außen als auch der Zellkern in der Zelle sind von einer
biologischen Membran umhüllt. Damit die Erbinformation
genutzt werden kann, wird von dem Teil, der gerade benötigt
wird, eine Kopie angelegt. Diese wandert dann ins Plasma,
wird dort verarbeitet und dann wieder zerstört. Jede Zelle
enthält die gesamte Erbinformation, es wir eben nur die
benutzt, die eine bestimmte Zelle gerade braucht. So wird
beispielsweise in einer Haarbalgzelle neben einigen anderen
der Teil des Genoms abgelesen oder kopiert, der etwas über
die Haarlänge und die Bildung und Verteilung der
Farbpigmente aussagt. Für eine Muskelzelle dagegen ist nur
interessant wie der Bauplan für eine Muskelfaser aussieht
und mit welchen Stoffwechselvorgängen eine Kontraktion
zustande gebracht werden kann.
Chromosomen im Transportformat
Die ganze Sache ist ziemlich kompliziert und
wir wollen das auch nicht weiter vertiefen. Wir merken uns
nur, daß jede Zelle das gesamte Genom enthält, daß jedoch
lediglich der Teil der Information genutzt wird, der für
eine bestimmte Leistung einer bestimmten Zelle gebraucht
wird, und daß von diesem Teil des Genoms eine Kopie
angefertigt wird, die dann zur Verarbeitung durch die
Kernmembran hindurch ins Zellplasma geschleust wird. Damit
die Kopien schnell und fehlerlos hergestellt werden können,
braucht man viel Platz. Daher sind die Chromosomen in diesem
Stadium ganz langgestreckt. Wenn man sich so eine Zelle im
Mikroskop anschaut, sieht man nur einen Wirrwarr von ganz
feinen Fäden im Zellkern, einzelne Chromosomen kann man
nicht erkennen. Wenn sich nun so eine Zelle teilen soll, sei
es, um eine verbrauchte oder verletzte Zelle zu ersetzen,
oder von der befruchteten Eizelle ausgehend einen ganz neuen
Organismus aufzubauen, dann muß gewährleistet sein, daß die
beiden neu entstehenden Tochterzellen genau das
gleiche Genom erhalten wie die Ausgangszelle. Dazu werden
die Chromosomen in der Vorbereitung einer Teilung oder
Mitose in eine Transportform gebracht. Sie werden nach
bestimmten Regeln aufgewickelt (spiralisiert) und mit
Proteinen verpackt. Am Ende dieser sogenannten Prophase
sind die Chromosomen kurz, kompakt und im Zellkern deutlich
sichtbar. In der nun folgenden Metaphase ordnen sich
die Chromosmen in der Äquatorialebene an, einer
gedachten Ebene, die den kugeligen Zellkern am Äquator
durchschneidet. Gleichzeitig löst sich die Kernmembran auf.
Jetzt kann man die Chromosomen im Mikroskop deutlich sehen
und voneinander unterscheiden. Wir sehen in Abb. 1 rechts
(aus R. Wolff, 1970, Katzen, S. 192), daß die Zellen der
Katze 38 Chromosomen haben.
Wenn wir aber noch genauer hinschauen und uns
die Mühe machen, die Chromosomen zu sortieren (Abb. 1
links), dann können wir noch viel mehr erkennen. Zu
jedem Chromosom gibt es einen Zwilling, der identisch
aussieht. Man nennt solche gleich aussehenden Chromosomen
auch homologe Chromosomen. Es gibt also 19
Chromosomenpaare oder Homologenpaare. Und jedes
einzelne Chromosom besteht aus zwei Spalthälften, den
Chromatiden. Was hat es nun damit auf sich?
Chromatiden lassen sich
vereinfach darstellen
Um das herauszukriegen, basteln wir uns
wieder, wie schon im ersten Teil, ein ganz einfaches
Modell. Da die Arbeit mit 38 Chromosomen nur zu einem
heillosen Durcheinander führen würde, tun wir so, als ob die
Katze nur 6 Chromosomen hätte. Dann sieht die
Äqatorialplatte der Metaphase wie in Abb. 2
schematisch dargestellt aus.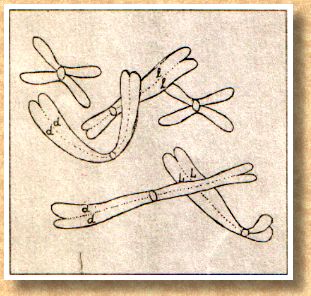
Jedes Chromosom hat eine "Kupplung", an der
später eine Art "Zugseil", die Spindelfaser,
angeheftet wird, mit deren Hilfe die Chromosomen auf die
beiden Tochterzellen verteilt werden. Diese Stelle wird
Zentromer genannt und ist im Schema durch einen Kreis
gekennzeichnet. Die Lage des Zentromers ist bei bestimmten
Chromosom immer gleich, entweder mehr in der Mitte, zu einem
Ende hin verschoben oder fast ganz an einem Ende. Außerdem
sehen wir in Abb. 1 und Abb. 2, daß die
Chromosomen unterschiedlich groß sein können. Jetzt haben
wir schon zwei Unterscheidungsmerkmale, mit deren Hilfe wir
Homologenpaare suchen können. Es ergibt sich zwangsläufig
die PaarAnordnung wie in Abb. 3 mit drei
Homologenpaaren.
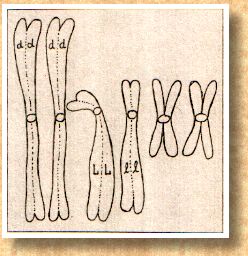 Was
sollen die Buchstaben auf zwei von den drei Homologenpaaren?
Wir wissen inzwischen, daß die Chromosomen die Träger der
Gene sind und daß die Abfolge der Nucleotide auf dem
DNS-Strang die Information für ein Merkmal codiert. Mit
unserer bereits genetisch geübten Denkweise schließen wir,
daß homologe Chromosomen so gleich aussehen, weil sie auch
gleiche Gene tragen, und liegen damit absolut richtig.
Allerdings können die beiden sich entsprechenden Gene auf
den homologen Chromosomen durchaus verschiedene Allele
codieren. Was
sollen die Buchstaben auf zwei von den drei Homologenpaaren?
Wir wissen inzwischen, daß die Chromosomen die Träger der
Gene sind und daß die Abfolge der Nucleotide auf dem
DNS-Strang die Information für ein Merkmal codiert. Mit
unserer bereits genetisch geübten Denkweise schließen wir,
daß homologe Chromosomen so gleich aussehen, weil sie auch
gleiche Gene tragen, und liegen damit absolut richtig.
Allerdings können die beiden sich entsprechenden Gene auf
den homologen Chromosomen durchaus verschiedene Allele
codieren.
Gene werden mit englischen
Begriffen bezeichnet
Man ist nun übereingekommen, daß man die Gene mit den
Anfangsbuchstaben der englischen Worte für das betreffende
Merkmal bezeichnet. So steht d für dilution
=Verdünnung, also für das Gen, das die Farbdichte bestimmt
und zugleich für das Allel der verdünnten Farbe, im Falle
einer schwarzen Katze also für blau. Trägt die Katze
das Allel D, dann zeigt ihr Fell die unverdünnte
Farbe, sie ist schwarz. Die Groß- und Kleinschreibung
drückt aus, in welcher Beziehung die Allele eines Gens
zueinander stehen. Das großgeschriebene dominante
oder beherrschende Allel setzt sich in jedem Fall durch,
egal wie das zweite Allel eines Genpaares auf dem homologen
Chromosom aussieht. Befindet sich dort das kleingeschriebene
rezessive oder zurückweichende Allel, hat dies keinen
Einfluß auf die Merkmalsausprägung. Eine Katze ist schwarz,
wenn das Genpaar für Farbdichte auf beiden homologen
Chromosomen das dominante Allel D trägt. Sie ist aber auch
dann schwarz, wenn nur eines der Homologen das Allel D
aufweist, das rezessive d auf dem anderen homologen
Chromosom spielt keine Rolle. Ist aber auf den beiden
Homologen das Allel d vorhanden, dann wird die
Farbverdünnung "eingeschaltet", die Katze ist blau.
Rezessive Allele auf beiden Homologen prägen den Phänotyp
Ebenso steht l für longhair =Langhaar und
damit für das Gen, das die Haarlänge bestimmt. Nur wenn auf
beiden Homologen das rezessive l zu finden ist, trägt
die Katze ein langes Haarkleid. Kurzhaarkatzen haben zweimal
das dominante L oder auf dem einen Chromosom L
und auf dem dazu homologen das rezessive l. Um sich
die ganze umständliche Beschreibung zu ersparen, spricht man
einerseits vom Phänotyp und bezeichnet damit das
äußere Erscheinungsbild bezüglich des zu beobachtenden
Merkmals. Dem stellt man den dazugehörigen Genotyp
gegenüber und schreibt die betreffenden Allelenpaare, soweit
bekannt, durch Schrägstrich voneinander getrennt
hintereinander. So hat die phänotypisch blaue Katze den
Genotyp d/d, etwas anderes ist nach dem oben gesagten
gar nicht möglich, denn d ist das rezessive Allel.
Wenn beide Allele eines Genpaares gleich sind, dann
bezeichnet man den Genotyp auch als homozygot. Woher
dieser Fachausdruck stammt, werden wir später noch sehen,
wenn wir nach der Meiose die Befruchtung besprechen, nehmen
wir den Begriff erst mal einfach so hin. Für die
phänotypisch schwarze Katze sind dagegen zwei Genotypen
denkbar, nämlich D/D oder D/d. Das Allel für
die unverdünnte Farbe muß nur mindestens einmal vorkommen.
Wenn, wie hier im zweiten Fall, die Allele eines Genpaares
verschieden sind, dann spricht man von einem
heterozygoten Genotyp. Man kann daher einer schwarzen
Katze den Genotyp nicht ohne weiteres ansehen, man schreibt
dann für das zweite, unsichere Allel einen Bindestrich als
Platzhalter, bis man vielleicht durch gründliche
Stammbaumanalyse oder durch die Nachkommen aus einer
bestimmten Verpaarung das zweite Allel enttarnt und
herausbekommt, ob die Katze bezüglich der unverdünnten Farbe
homozygot oder heterozygot ist. Der Genotyp D/- weist
also in unserem Fall auch auf eine phänotypisch schwarze
Katze hin. Die Kurzhaarkatze ist genotypisch entweder L/L,
L/l oder L/-. Langhaarige Katzen müssen
dagegen homozygot mit l/l ausgestattet sein. Und die
Halblanghaar? Das ist ein Fall der zeigt, daß kaum ein Gen
für sich alleine für ein Merkmal verantwortlich ist. Häufig
spielen neben dem Hauptgen noch ganz andere Gene eine Rolle
bei der Ausprägung eines Merkmals. Die Haarlänge ist ein
solches polygen veranlagtes Merkmal mit einer großen
Variationsbreite, denn die halblanghaarigen mit l/l
sind ebenso homozygot wie die langhaarigen.
Das
dritte Chromosomenpaar lässt sich finden
Jetzt wissen Sie sicher, was die Buchstaben
auf den Chromosomen in Abb. 2 und Abb. 3 zu
bedeuten haben. Ich habe für unser Modell nicht nur die
Chromosomenzahl reduziert, sondern ich will sogar noch einen
Schritt weitergehen. Nehmen wir jetzt und für unsere
zukünftige Arbeit an, daß man auf den Chromosomen nicht nur
bestimmte Gene lokalisieren kann, sondern sogar sehen kann,
welches Allel auf welchem Chromosom sitzt. Da unsere
ModellKatze von der Grundfarbe her schwarz sein soll, sorgt
das homozygote d/d auf dem langen 1. Chromosomenpaar
mit dem Zentromer in der Mitte dafür, daß sie phänotypisch
blau aussieht. Mit dem heterozygoten L/l auf dem
kürzeren 2. Chromosomenpaar mit dem aus der Mitte
verschobenen Zentromer muß sie kurzhaarig sein. Und was ist
mit dem 3. Chromosomenpaar ohne Buchstaben? Überlegen Sie
mal selber, vielleicht kommen Sie darauf, wenn Sie darüber
nachdenken, ob es sich um eine Katze oder einen Kater
handeln könnte.
Zugegeben, wir sind mit unserem Programm
nicht so weit gekommen, wie geplant. Mit der Mitose haben
wir gerade eben angefangen, von der Meiose noch nichts
gehört. Aber diese Vorüberlegungen haben sich so
selbstverständlich aus der allgemeinen Einführung in die
Mechanismen der Zellteilungen ergeben, daß sie nicht
ignoriert werden konnten. Mit dem jetzt angesammelten Wissen
führen wir das nächste Mal die Mitose schnell zu Ende und
vertiefen uns in die Meiose. Dabei lösen sich auch die
Rätsel "3. Chromosomenpaar" und "Chromatiden" fast von
alleine.
mit freundlicher Genehmigung des Autors
Dipl. Biologe R.
Fahlisch
"Dreamhunter Cattery"
Das Copyright für den oben genannten Text, liegt sowohl beim
Autor des Textes Herrn R. Fahlisch, sowie bei dem Betreiber
dieser Seiten, Frau Ute Kunze. Eine Vervielfältigung oder
Verwendung des Textes in anderen elektronischen oder
gedruckten Publikationen ist ohne ausdrückliche Zustimmung
von Herrn Fahlisch und Frau Kunze nicht gestattet. |
|